Половой гормон у кошек
К ВОПРОСУ О ВОЗМОЖНОСТИ РАЦИОНАЛЬНОЙ ГОРМОНАЛЬНОЙ СУПРЕССИИ ПОЛОВОЙ ФУНКЦИИ КОШЕК.
В.М. Брохин,
Ассоциация врачей ветеринарной медицины, ФОНД РАЗВИТИЯ ВЕТЕРИНАРИИ, СПб, пейджер (812) 163-50-56 абонент 680
Последние несколько лет многие коллеги обратили внимание на значительное увеличение в практике случаев кистозной гиперплазии эндометрия у кошек. По данным, собранным различными врачами, у этих пациентов в анамнезе почти всегда выявляется некорректное применение гормональных препаратов для супрессии половой функции. К сожалению, страсть владельцев животных к фармакологическим экспериментам неистребима. Однако и врачам есть о чем задуматься. В доступной литературе вопросы акушерства и гинекологии кошек раскрыты недостаточно. Изредка же можно наткнуться на вовсе шокирующие сведения. Например в книге Я.Г. Губаревича «Акушерство мелких животных», М.,Л.,1952г., читаем: «У кошек течка наблюдается два раза в год в конце зимы и в конце лета.» И там же: «Движения полового члена во влагалище причиняют кошке боли, вследствие чего она во время совокупления издает своеобразный писк. Болезненность у кошек во время полового акта объясняется тем, что на головке полового члена кота имеются роговые зубчики» [5]. От комментариев воздержимся!
В связи с вышеизложенным представляет интерес рассмотреть современные данные по физиологии [1,3], что даст возможность определить наиболее рациональные подходы к применению гормональных препаратов для супрессии половой функции кошек.
Нормальная физиология полового цикла кошки
Половое созревание кошек наступает в возрасте 4-12 месяцев. Репродуктивный период длится 8-10 лет. Для кошек характерна сезонная полиэструсность в течение периода, когда световой день не менее 12-14 часов (с января по сентябрь для северного полушария). Овуляция индуцируется коитальным шеечно-влагалищным рефлексом, который может быть вызван и симулированным коитусом. Эструс насту-пает каждые 4-36 дней (в среднем 14-30).
Проэструс длится от нескольких часов до двух дней, в течение которых выделяются феромоны. Отмечается также небольшое выделение мукоидного секрета бартолиевых желез. Эструс обычно длится 6-10 дней, однако возможны колебания от 2 до 12. Продолжительность эструса не зависит от овуляции и не связана с ней. Овуляция происходит через 24-50 часов после копуляции (в среднем 24-30) в ходе реализации шеечно-влагалищного рефлекса. Сперма в матке остаётся достаточно активной в течении 24 часов(от 2 до 24), а значит оплодотворение наступает в период до 48 часов после овуляции. Зигота движется по яйцеводу 4 дня. Имплантация происходит на 14 день после оплодотворения, после чего формируется зонарная эндотелио-хориональная плацента. Период гестации 58-70 дней (обычно60-63).
Диэструс. Вслед за псевдокопуляцией с последующей овуляцией образует-ся желтое тело, которое функционирует 30-40 дней. В этом случае весь цикл длится около 6 недель.
Постэструс наступает, если у кошки не было овуляции или опло-дотворения. Эта стадия длится 7-21 дней, затем яйцеклетка дегене-рирует и половой цикл возобновляется с проэструса.
Анэструс длится от 1 до 6 месяцев, в зависимости от длины светового дня. В этот период кошки не спариваются.
Определение фазы полового цикла по вагинальным мазкам
Любое применение половых гормонов требует аккуратного предварительного изучения гормонального статуса пациента. Определение содержания гормонов в крови на сегодня малодоступно, поэтому остается более распространенный метод определения фаз полового цикла и гормонального статуса по влагалищным мазкам [2]. Вагинальный мазок для цитологического исследования может быть получен с помощью 3-х мм влажного тампона, введенного во влагалище на 1,5-2см. В некоторых случаях можно инстиллировать, а затем аспирировать глазной пипеткой несколько капель стерильного физиологического раствора. Здесь необходимо помнить, что применение пипетки может симулировать коитус и вызвать овуляцию.
После обычной фиксации на предметном стекле мазок окрашивается метиленовой синью или по Романовскому. Важно также отметить, что у кошек в процессе полового цикла не наблюдается диапедез эритроцитов во влагалище. При цитологическом исследовании влагалищных мазков обнаруживаем:
АНЭСТРУС. Редкие фрагменты клеток, лейкоциты; выраженное количество мелких эпителиальных клеток с высоким соотношением ядро/цитоплаз-ма, с сентября по январь такие клетки часто встречаются в группах.
ПРОЭСТРУС. Увеличивается число фрагментов; появляются редкие, но крупные эпителиальные клетки с низким соотношением ядро/цитоплазма; могут быть отмечены единичные лейкоциты.
ЭСТРУС. Значительно падает число фрагментов; выраженное количество многолучевых клеток с извилистыми краями; ядра этих клеток темные, точечные или отсутствуют; лейкоциты отсутствуют.
РАННИЙ ДИЭСТРУС. Неясные, оборванные края многолучевых клеток; практически не обнаруживаются лейкоциты, появляются бактерии и увеличивается число фрагментов.
ПОЗДНИЙ ДИЭСТРУС. Увеличивается число мелких базофильных эпителиальных клеток; лейкоциты все еще присутствуют (весь период метэструса 7-21 день); если овуляции не было, мазок покажет возврат к анэструсу с возможным присутствием единичных лейкоцитов.
Возможности гормональной профилактики нежелательной беременности.
- Эстрогены, назначенные в период проэструса и эструса или только эструса курсом до 10-14 дней, предотвратят развитие маточного эпителия и, удлинив эструс, заблокируют имплантацию зиготы. Кроме того, поддержание уровня эстрогенов в крови предотвратит образование в половых путях среды, необходимой для оплодотворения.
- Комбинированное назначение эстрогенов и гестагенов заблокирует овуляцию, если их назначить курсом на период проэструса и эструса.
- Назначение гестагенов во вторую половину анэструса или в лютеальную фазу диэструса позволит симулировать функционирование желтого тела беременности и животное не придет в охоту.
Если ставится задача предпринять какие либо меры после нежелательной вязки, возможно применение эстрогенов. Тем не менее, необходимо учитывать вполне вероятную возможность развития эндометрита или кистозной гиперплазии матки [4].
При использовании комбинированных препаратов, мы даем кошке полную свободу и заботимся о соблюдении регулярности в приеме таблеток. Метод требует значительных усилий, постоянного наблюдения за животным и удовлетворяет скорее кошку, но не ее хозяев. Метод подходит скорее для сук, так как они приходят в охоту 1-2 раза в год и проведение назначенного курса препарата не слишком затруднит владельцев.
Если владельцы просят врача прекратить мешающие им проявления функционирования половой системы кошки, но отказываются от кастрации, лучше всего назначить гестагены пролонгированного действия, например, хорошо известный медроксипрогестерона ацетат (Неонидан®, КРКА) и повторять его инъекции каждые 3-4 месяца в течение всего сезона. В этом случае врач должен помнить, что характерных для всех половых гормонов побочных эффектов нужно избегать. Для этого и используют метод вагинальных мазков [2,3], позволяющий определить гормональный фон организма. Желательной для начала курса инъекций лютеальной фазы полового цикла можно достичь искусственно, применив в период эструса симулированный коитус и добившись таким образом овуляции с последующим диэструсом.
Источник
Половой гормон у кошек
Особенности физиологии и эндокринологии кошек
Дж. Верстеген (John Verstegen)
За последнее десятилетие была создана внушительная база данных, касающихся взаимосвязи поведенческих, яичниковых и эндокринных изменений, происходящих в организме кошек в течение анэструса, эструса и лютеиновой фазы после стерильной или фертильной вязки, во время беременности и в послеродовой период. Однако в знаниях о репродуктивной физиологии и эндокринологии все еще существуют пробелы и противоречия, требующие дальнейших исследований.
Эстральный цикл у кошек имеет сезонный характер (хотя бывают исключения) и во многом сходен с циклом таких млекопитающих, как кролики, норки и хорьки, особенно в том, что касается индукции овуляции.
Средний возраст полового созревания у домашней кошки варьирует. Как правило, первый эструс начинается, когда вес животного достигает 2,3–2,5 кг. В норме самка может достичь половой зрелости не раньше, чем в 4 месяца и не позднее, чем в 18 месяцев, но в большинстве случаев первые признаки сексуальной активности проявляются в 6–9 месяцев.
В какой-то степени возраст полового созревания определяется наследственностью, некоторые породы демонстрируют раннее или, наоборот, относительно позднее созревание. Наиболее «скороспелыми» являются короткошерстные породы (сиамская, бирманская), животные достигают зрелости при меньшем весе, чем кошки длинношерстных пород (персидская), у которых первая течка может начаться в возрасте 18 месяцев. Однако насколько можно судить, основным фактором, определяющим сроки полового созревания и установление цикла, являются длительность и интенсивность светового дня или искусственного освещения (для животных, содержащихся в помещении).
Достижение половой зрелости зависит от времени года и, как правило, происходит в то время, когда длительность светового дня увеличивается. Самки, вступающие в пубертатный период в конце лета, на фоне продолжительного светового дня демонстрируют поведенческий эструс в более раннем возрасте, чем те, что достигают зрелости осенью или зимой, — в последнем случае течка наблюдается лишь следующей весной.
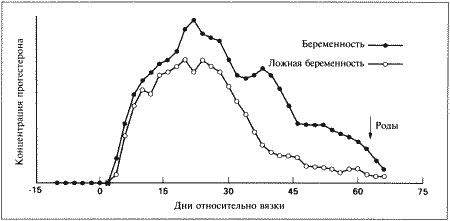 Фиг. 2.1.
Фиг. 2.1.
Изменения концентрации прогестерона в плазме крови при физиологической и ложной беременности. Воспроизведено из кн. Lane и Cooper «Veterinary Nursing», (1994) с разрешения Butterworth Heinemann
СЕЗОННЫЙ ХАРАКТЕР ЭСТРАЛЬНОГО ЦИКЛА
В отсутствие беременности или при ложной беременности эструс у кошек возобновляется каждые 2–3 недели весной, летом и осенью. При содержании в помещении с искусственным освещением, особенно если свет горит и ночью, периодичность цикла может проявляться вне зависимости от времени года, в таких случаях течка у кошек наблюдается и зимой. Влияние освещения на репродуктивный цикл осуществляется через эпифиз и вырабатываемый им гормон мелатонин. Механизм воздействия, видимо, сходен с тем, что наблюдается у кобыл.
С точки зрения поведения цикл кошки можно условно разделить на период течки и период ее отсутствия.
Проэструс и эструс
Период течки разделяют на проэструс и эструс. Поведенческие проявления проэструса и эструса наблюдаются в течение 3–10 дней. Переход от проэструса к эструсу у кошек в отличие от собак не выражен, поскольку за исключением поведенческих обе фазы не характеризуются заметными внешними проявлениями. Эструсом называют период, когда кошка испытывает половое возбуждение и подпускает к себе самца. Первые дни течки (с 1 по 4 день), на протяжении которых самка демонстрирует поведенческий эструс, но не подпускает к себе самца, называют проэструсом. Определить фазу проэструса с применением клинических методов затруднительно, поскольку некоторые самки подпускают к себе самца одновременно с наступлением поведенческого эструса, т. е. минуя подготовительный период проэструса. Развитие фолликулов в период проэструса начинается на фоне сниженной концентрации эстрадиола, недостаточной для обеспечения максимальных проявлений поведенческого эструса.
Начало эструса определяют по тому, что кошка подпускает к себе самца, а о его завершении судят по окончанию таких поведенческих проявлений. Период эструса характеризуется максимальным уровнем эстрогенов, вырабатываемых фолликулами.
В период течки кошка трется о людей и предметы, катается, зачастую проявляет крайнее дружелюбие. Характерным признаком являются протяжные призывные крики. При поглаживании, особенно по спине, кошка принимает характерную позу — выгибает поясницу, приподнимает круп, отводит в сторону хвост и перебирает лапами. Самцы проявляют интерес к самке, которая, вступив в период течки, может убежать из дома для случки. Во время вязки кошка издает громкие крики и с силой, а иногда и агрессивно пытается освободиться от самца, удерживающего ее за холку. После вязки половое возбуждение на некоторое время утихает; однако вскоре за первой следуют новые, многократные копуляции, как правило, наблюдаемые на протяжении всего эструса. Существует предположение, что вязки сокращают период эструса, хотя проверить справедливость этого утверждения в условиях контролируемого разведения достаточно сложно.
Осмотр наружных половых органов не позволяет выявить изменений, однозначно свидетельствующих о наступлении эструса. В некоторых случаях наблюдается небольшое покраснение или припухание вульвы, однако выделения отсутствуют. Иногда становится видна половая щель, незаметная в период анэструса.
Интерэструс и анэструс
При отсутствии вязки и/или спонтанной овуляции течка возобновляется каждые 10–14 дней в течение всего репродуктивного сезона. Интервал между двумя периодами интенсивной сексуальной активности, во время которого у животного не наблюдается специфических физиологических и поведенческих признаков, называют интерэструсом. Интерэструс характеризуется относительным покоем яичников и матки, связанным с подготовкой к очередному циклу развития фолликулов и соответственно следующему эструсу. В период интерэструса концентрация эстрогенов снижается до базального уровня.
Однако у некоторых животных циклы развития фолликулов следуют один за другим без промежутков. В этом случае концентрация эстрогенов не снижается, и кошка может демонстрировать постоянный эструс. Такое явление часто ошибочно называют нимфоманией или затянувшимся эструсом.
В период анэструса, который характеризуется полным покоем яичников, в отличие от интерэструса наблюдается длительное отсутствие как сексуальной активности, так и признаков репродуктивного поведения. Начало анэструса совпадает с сокращением светового дня или искусственного освещения до 4–6 часов в сутки.
Как правило, у животных северного полушария анэструс наблюдается поздней осенью или в начале зимы, но его может и не быть, если кошка живет в условиях светового дня постоянной длительности.
ВЯЗКА И ИНДУЦИРОВАННАЯ ОВУЛЯЦИЯ
Говоря об особенностях репродукции кошки, как правило, отмечают индуцированный характер овуляции, т. е. овуляция наблюдается как реакция на коитус (естественная — в процессе копуляции, или искусственная — в результате вагинальных манипуляций), хотя в некоторых случаях она может происходить спонтанно. Вагинальная стимуляция во время копуляции вызывает возбуждение нервных волокон, передающееся в гипоталамус и служащее пусковым механизмом для выброса ГнРГ. Выброс ГнРГ в свою очередь стимулирует выделение лютеинизирующего гормона ЛГ. Аналогичным образом введение экзогенных гонадотропинов или ГнРГ вызывает овуляцию нескольких яйцеклеток в течение 24–48 часов после введения. Скорость овуляции, очевидно, напрямую связана с амплитудой пика ЛГ, зависящей от количества и интенсивности вязок. Интервал между вагинальной стимуляцией и овуляцией обратно пропорционален количеству вязок и эндокринному статусу при копуляции. При вязке, состоявшейся в середине эструса, овуляция происходит раньше, чем при вязке, имеющей место в начале эструса. Но в любом случае овуляция происходит не ранее, чем через 24, и не позднее, чем через 52 часа после индуцированного выброса ЛГ. Через 24–48 часов после овуляции течка прекращается. Однако в некоторых случаях выброс ЛГ происходит спонтанно — в отсутствие вязки или вагинальной стимуляции, но также вызывает овуляцию. Характерное повышение концентрации прогестерона, наблюдающееся после овуляции, зафиксировано у кошек, не участвовавших в вязке. Спонтанная овуляция чаще наблюдается у старых животных либо у кошек в питомниках, если они содержатся в одном помещении с котами, причем даже в отсутствие визуального и физического контакта. Вероятно, данное явление объясняется воздействием феромонов.
Поведенческие аспекты вязки и возможные проблемы рассматриваются в гл. 10.
ГОРМОНАЛЬНАЯ ПОДДЕРЖКА ОВУЛЯЦИИ, ЛОЖНАЯ БЕРЕМЕННОСТЬ И БЕРЕМЕННОСТЬ
Принято считать, что у кошек, так же, как и у других животных, развитие фолликулов обеспечивается воздействием фолликулостимулирующего гормона (ФСГ), а клинические и поведенческие изменения, связанные с периодами проэструса и эструса, обусловливаются выработкой эстрогенов. Однако в настоящее время не имеется достоверных данных об изменении концентрации ФСГ в плазме за время эстрального цикла. Если овуляция не происходит, фолликулы атрофируются, концентрация эстрогенов падает. Лютеиновая фаза не наступает, и после нескольких дней интерэструса начинается очередная волна роста фолликулов.
Индуцированный или спонтанный выброс ЛГ стимулирует овуляцию и развитие желтых тел. Как правило, самка подпускает самца на второй или третий день после начала роста фолликулов (1 и 2 день эструса), однако у некоторых животных всплеск ЛГ как реакция на копуляцию наблюдается лишь на 4 или 5 день. Видимо, предварительное (за несколько дней до копуляции) повышение концентрации эстрогена является необходимым условием для выброса ЛГ и овуляции. Дальнейшее развитие репродуктивного цикла у кошек в отличие от собак зависит от фертильности вязки.
Инфертильная вязка (без оплодотворения)
При отсутствии оплодотворения яйцеклеток желтые тела развиваются и вырабатывают прогестерон в течение приблизительно 25–45 дней. Эта лютеиновая фаза короче аналогичного периода, наблюдаемого при развитии беременности, потому ее часто называют ложной беременностью. Таким образом, ложная беременность у кошек представляет собой лютеиновую фазу, следующую за овуляцией, которая не завершилась оплодотворением и имплантацией, однако у них — в отличие от собак — не бывает изменений в поведении и лактации. В течение этого периода секреция ГнРГ угнетена, что обеспечивает высокий уровень прогестерона, препятствующий возобновлению течки. Лютеиновая фаза, или ложная беременность завершается коротким интерэструсом, предшествующим возобновлению эструса, если продолжается брачный сезон или наступает фаза анэструса.
Овуляция и оплодотворение
После вязки с фертильным самцом или успешного искусственного осеменения эмбрионы в течение 4–5 дней после оплодотворения развиваются в маточных трубах, затем мигрируют в рог матки, где приблизительно на 12–16 день после повторной вязки (первая необходима для индукции овуляции) происходит имплантация. Перед имплантацией бластоцисты свободно перемещаются в полости матки, мигрируя из одного рога в другой, вероятно, такая миграция обеспечивает равномерное распределение эмбрионов в полости матки. Считая от момента имплантации, беременность продолжается еще 50 дней, таким образом, общая продолжительность беременности составляет около 64 дней (от даты первой вязки). Секреция прогестерона во время беременности — как и в случае ложной беременности — препятствует наступлению эструса.
После окончания лактации, если брачный сезон продолжается, — наблюдается короткий период интерэструса, сменяющийся эструсом; если брачный сезон закончен — наступает анэструс. Как правило, эструс возобновляется через 10–15 дней после прекращения кормления. Однако иногда течка может возникать у кормящих кошек спустя 10–15 дней после родов. В таких случаях, если не произошла овуляция, — нормальный эструс возобновляется каждые 10–20 дней. Первая вязка после родов часто не приводит к оплодотворению в связи с неполной инволюцией матки. Зато вязки в период следующего эструса бывают фертильными, поэтому нередко можно наблюдать кормящую кошку, беременную новым пометом. Имеются данные о том, что в некоторых случаях во время беременности у кошек наблюдается спонтанный или индуцированный рост фолликулов, сопровождающийся поведенческим эструсом и вязками. Данные факты позволяют предположить, что и в период беременности яичники кошки чувствительны к воздействию гонадотропинов.
Развитие фолликулов сопровождается секрецией 17-?-эстрадиола, концентрация которого достигает максимального уровня в период эструса. В течение анэструса и интерэструса концентрация эстрадиола поддерживается на базальном уровне ( 100 нг/мл после максимальной стимуляции. Оптимальный выброс ЛГ наблюдается после 4 случек, происходящих в течение 2–4 часов. Дальнейшее увеличение интенсивности или продолжительности коитальной стимуляции не оказывает существенного влияния на повышение концентрации ЛГ, что, очевидно, объясняется истощением гипофиза.
В период анэструса, интерэструса, проэструса и эструса концентрация прогестерона поддерживается на базальном уровне вплоть до овуляции.
У кошек как при беременности, так и в ее отсутствие — концентрация прогестерона в плазме начинает повышаться после овуляции, через 24–50 часов после выброса ЛГ. Максимальные показатели прогестерона (100–200 нмоль/л) наблюдаются на 20–25 день после первой вязки.
При ложной беременности концентрация прогестерона начинает снижаться приблизительно на 25 день и достигает базального уровня на 30–40 день. Такое постепенное снижение концентрации прогестерона является характерной особенностью ложной беременности у кошек. При истинной (физиологической) беременности концентрация прогестерона поддерживается на высоком уровне в течение всего периода вынашивания и стремительно снижается перед родами. Постепенное снижение концентрации прогестерона в конце ложной беременности у кошек сходно с тем, что наблюдается у собак, и, вероятно, объясняется отсутствием лютеолитического фактора, вырабатываемого в организме самок в конце беременности. Очевидно, на 25–35 день ложной беременности желтые тела, лишенные лютеотрофической поддержки со стороны эмбрионов и плаценты, подвергаются «запрограммированной» атрофии. Гистероэктомия, выполненная во время лютеиновой фазы ложной беременности, не отражается на жизнеспособности желтых тел, что свидетельствует о том, что матка не участвует в процессе рассасывания желтых тел.
Снижение концентрации прогестерона начинается на 25–35 день беременности, после чего вплоть до 60 дня ее значения поддерживаются на уровне 15–30 нмоль/л (5–10 нг/мл). Прогестерон необходим кошкам в течение всего срока вынашивания для сохранения беременности, хотя беременность может сохраняться в течение нескольких дней после того, как концентрация прогестерона в плазме крови снижается Вагинальные выделения кошки (а). Выделения, типичные для фазы эструса. Большая часть клеток представляет собой безъядерные кератинизированные клетки или клетки с пикнотическим ядром. Присутствуют промежуточные клетки (b). Выделения, характерные для метэструса, — «течки», иногда наблюдаемой у кошек в конце эструса. Присутствуют поверхностные и промежуточные клетки, повышено содержание лейкоцитов. Этот короткий метэструс наблюдается в течение 24–48 часов (см. Приложение)
Banks D. H. and Stabenfeld G. Y. (1982) Luteinizing hormone release in the cat it response to coitus on consecutive days of estrus. Biology of Reproduction 26, 603–611.
Burke T. J. (1976) Feline reproduction. Veterinary Clinics of North America 6, 317–321.
Concannon p, Hodson В. and Lein D. (19S0) Reflex L. H. release in estrous cats following single and multiple copula tions. Biology of Reproduction 23, 111–117.
Goodrowe K. L., Howard J. G. Schmidt P. M. and Wildt D. E. (1989) Reproductive biology of the domestic cat with special reference to endocrinology, sperm function and in-vitro fertilization. Journal of Reproduction and Fertility Supplement 39, 73–90.
Hurni H. (1981) Daylength and breeding in the domestic cat. Laboratory Animals 15, 229–233.
Olson P. N., Husted P. W., Allen Т. А. and Nett T. M. (1984) Reproductive endocrinology and physiology of the bitch and queen. Veterinary Clinics of North America: Small Animal Practice 14, 927–946.
Schmidt P. M., Chakraborty P. K. and Wildt D. E. (1983) Ovarian activity, circulating hormones and sexual behaviour in the cat II: relationships during pregancy, parturition, lactation and the postpartum estrus. Biology of Reproduction 28, 657–671.
Shille V. M., Lundstrom K. E. and Stabenfeldt G. H. (1979) Follicular function in the domestic cat as determined by estradiol 17 beta concentrations in plasma: relation to estrous behavior and cornifi-cation of exfoliated vaginal epithelium. Biology of Reproduction 21, 953–963.
Stewart D. R. and Stabenfeldt G. H. (1983) Relaxin activity in the pregnant cat. Biology of Reproduction 32, 848–854.
Tsutsui T. and Stabenfeldt G. H. (1993) Biology of ovarian cycles, pregnancy and pseudopregnancy in the domestic cat. Journal of Reproduction and Fertility Supplement 47, 29–35.
Verstegen J. P., Onclin K., Silva L. D. M., Wouters-Ballman P., Delahaut Fand Ectors F (1993) Regulation of progesterone during pregnancy in the cat: studies on the roles of corpor lutea, placenta and prolactin secretion. Journal of Reproduction and Fertility Supplement 47, 165–173.
Источник



